
 イネ科イネ亜科イネ族イネ属の多年生あるいは一年生草本で、穀物の一種。種子の外側には穎花の内外穎が変化した籾があり、その内側に玄米があります。イネ籾では長軸方向に内外穎の段差が明瞭にみられ、籾の表皮全体は顆粒状突起に覆われています。玄米の大部分は胚乳で占められ、小穂軸に近い外穎側の基部に胚が存在します。種子の基部には二枚の護穎があり、小穂軸によってイネの小穂梗につながっています。
イネ科イネ亜科イネ族イネ属の多年生あるいは一年生草本で、穀物の一種。種子の外側には穎花の内外穎が変化した籾があり、その内側に玄米があります。イネ籾では長軸方向に内外穎の段差が明瞭にみられ、籾の表皮全体は顆粒状突起に覆われています。玄米の大部分は胚乳で占められ、小穂軸に近い外穎側の基部に胚が存在します。種子の基部には二枚の護穎があり、小穂軸によってイネの小穂梗につながっています。
 アジアのイネは、考古学的証拠から約8000年以前の中国長江下流域で栽培化されたと考えられています。日本では以前、縄文時代のイネ・籾痕と報告された資料が少なからずありましたが、年代測定および再同定が行われた結果、後世の種子の混入や種実の同定に問題があり、現状ではイネの確実な資料は西日本の突帯文土器期(約2800年前)までしか遡りません(中沢2016)。
アジアのイネは、考古学的証拠から約8000年以前の中国長江下流域で栽培化されたと考えられています。日本では以前、縄文時代のイネ・籾痕と報告された資料が少なからずありましたが、年代測定および再同定が行われた結果、後世の種子の混入や種実の同定に問題があり、現状ではイネの確実な資料は西日本の突帯文土器期(約2800年前)までしか遡りません(中沢2016)。

- 参考文献
-
- 中沢道彦 2016「イネ・アワ・キビはいつ来てどのように栽培されたのか」『いま、アッコンが面白い!―タネ・ムシ圧痕が語る先史・古代の農とくらし―』日本学術振興会科学研究費補助金研究成果公開シンポジウム 熊本大学文学部小畑研究室
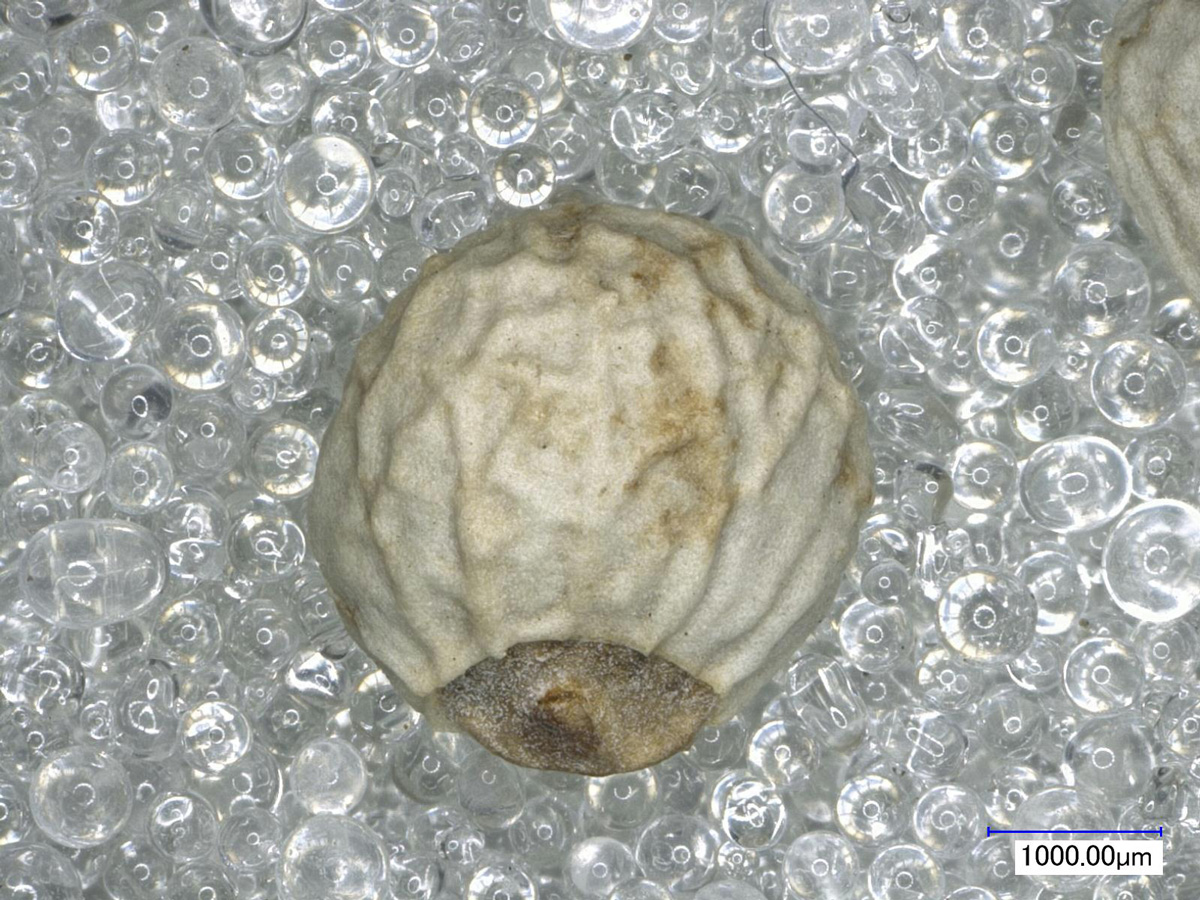
 イネ科ウシノケグサ亜科コムギ族オオムギ属の一年生草本で、穀物の一種。内外穎が穎果に癒着しているものを皮性オオムギ、容易に離れやすいものを裸性オオムギといいます。大きさのみでは両者を区別するのは難しいです。皮性オオムギは①裏面溝の上部が大きく開く、②断面形が六角形をなす、③脈線が顕著である、という特徴があります。一方で、裸性オオムギは①溝は幅広いものもあるが、その幅は上端部でも同じである、②断面系は角張らず滑らかな楕円形に近い、③脈線はほとんどない、という特徴を持ちます(小畑2011)。
イネ科ウシノケグサ亜科コムギ族オオムギ属の一年生草本で、穀物の一種。内外穎が穎果に癒着しているものを皮性オオムギ、容易に離れやすいものを裸性オオムギといいます。大きさのみでは両者を区別するのは難しいです。皮性オオムギは①裏面溝の上部が大きく開く、②断面形が六角形をなす、③脈線が顕著である、という特徴があります。一方で、裸性オオムギは①溝は幅広いものもあるが、その幅は上端部でも同じである、②断面系は角張らず滑らかな楕円形に近い、③脈線はほとんどない、という特徴を持ちます(小畑2011)。
 オオムギは中近東にて野生二条オオムギが栽培化され、六条種が遅れて出現しました。日本に最初に伝来したのは六条種とされています。日本列島での確実なオオムギの圧痕は、山梨県中道遺跡の縄文時代晩期か弥生時代前期となる土器に1点確認されているのみです(中沢・丑野ほか2002)。ただし、この資料も後代のものではないかとの説もあります。炭化資料では弥生時代後期の例が確認されています。
オオムギは中近東にて野生二条オオムギが栽培化され、六条種が遅れて出現しました。日本に最初に伝来したのは六条種とされています。日本列島での確実なオオムギの圧痕は、山梨県中道遺跡の縄文時代晩期か弥生時代前期となる土器に1点確認されているのみです(中沢・丑野ほか2002)。ただし、この資料も後代のものではないかとの説もあります。炭化資料では弥生時代後期の例が確認されています。
- 参考文献
-
- 小畑弘己 2011「オオムギ種子の形態学的観察」『東北アジア古民族植物学と縄文農耕』pp.209-219 同成社
- 中沢道彦・丑野毅・松谷暁子 2002「山梨県韮崎市中道遺跡出土の大麦圧痕土器について」『古代』第111号 pp.63-83 早稲田大学考古学研究会
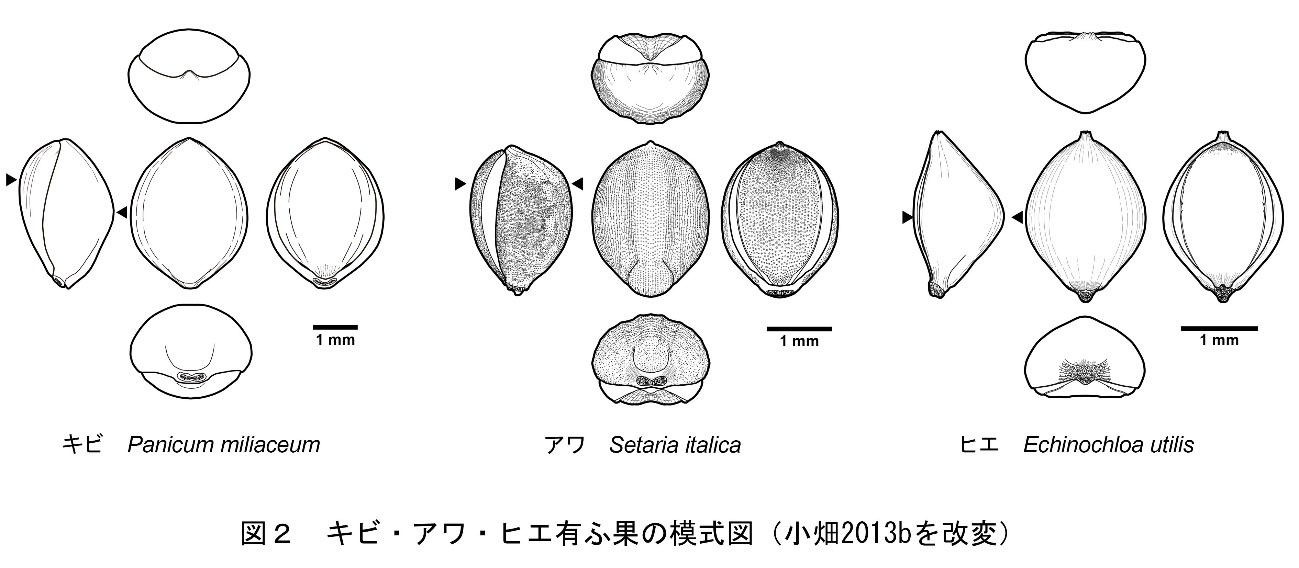
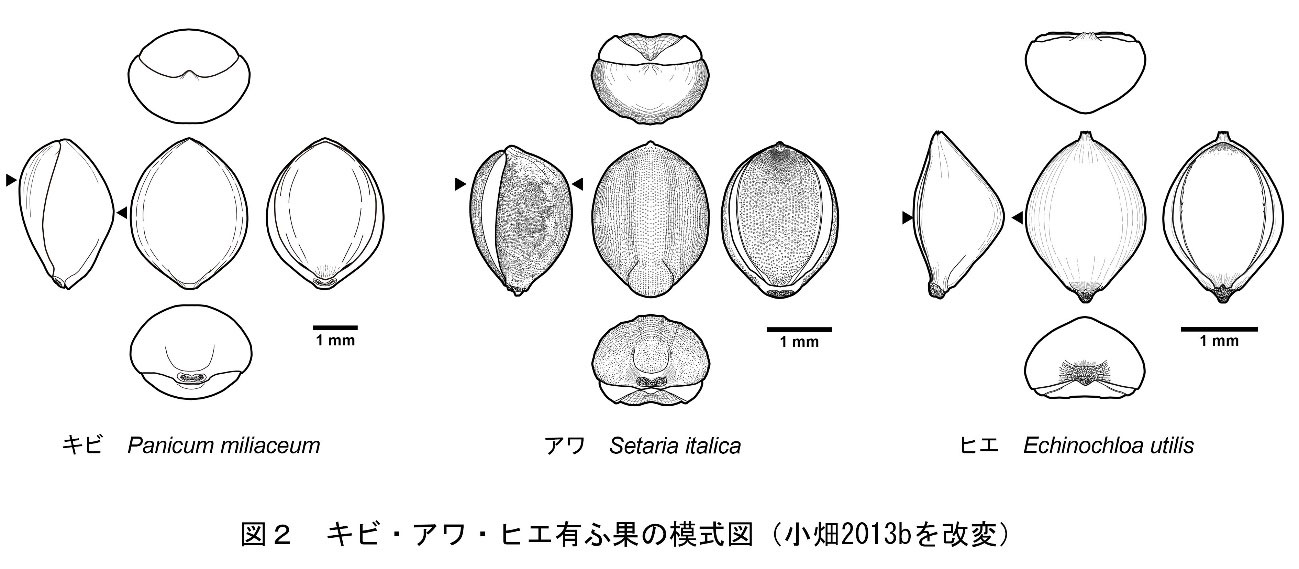
 イネ科キビ亜科キビ族アワ属の一年生草本で、穀物の一種。有粰果は平面観が丸みを帯びた紡錘形を呈し、基部側が台形状に突出します。内外穎の表面には乳頭状突起が存在し、内穎側は外穎と重なる部分が平滑な三日月となっています。内穎の中央部は溝状に窪み、先端部は点状に深く窪みます。側面部での最大部は、内外穎とも上位にあります。横断面形は内穎側が平坦な鈍角の隅丸五角形となります(小畑・真邉2014)。胚部は果実の長さの2/3を占め、幅も広いです。反対側の面には小さなヘラ形をした臍があります(椿坂1993)。
イネ科キビ亜科キビ族アワ属の一年生草本で、穀物の一種。有粰果は平面観が丸みを帯びた紡錘形を呈し、基部側が台形状に突出します。内外穎の表面には乳頭状突起が存在し、内穎側は外穎と重なる部分が平滑な三日月となっています。内穎の中央部は溝状に窪み、先端部は点状に深く窪みます。側面部での最大部は、内外穎とも上位にあります。横断面形は内穎側が平坦な鈍角の隅丸五角形となります(小畑・真邉2014)。胚部は果実の長さの2/3を占め、幅も広いです。反対側の面には小さなヘラ形をした臍があります(椿坂1993)。
 アワの原型はエノコログサ(Setaria viridis)と考えられており、栽培起源地には中国北部~内蒙古地域が想定されています。中国では約8000年前に栽培されていたことが明らかにされ、ロシア沿海州地方や韓国南部においては約5000年前にキビと共に栽培されています(小畑2011)。日本列島におけるアワの最古の証拠は、福岡県江辻遺跡および大分県石井入口遺跡から検出された約2800年前の突帯文土器出現期のアワ圧痕です(中沢2016)。
アワの原型はエノコログサ(Setaria viridis)と考えられており、栽培起源地には中国北部~内蒙古地域が想定されています。中国では約8000年前に栽培されていたことが明らかにされ、ロシア沿海州地方や韓国南部においては約5000年前にキビと共に栽培されています(小畑2011)。日本列島におけるアワの最古の証拠は、福岡県江辻遺跡および大分県石井入口遺跡から検出された約2800年前の突帯文土器出現期のアワ圧痕です(中沢2016)。

- 参考文献
-
- 小畑弘己 2011「縄文時代の栽培植物とその起源」『東北アジア古民族植物学と縄文農耕』pp.153-179 同成社
- 小畑弘己・真邉 彩 2014「韓国櫛文土器文化の土器圧痕と初期農耕」『国立歴史民俗博物館研究報告』第187集 pp.111-160
- 椿坂恭代 1993「アワ・ヒエ・キビの同定」『先史学と関連化学』吉崎晶一先生還暦記念論集 pp.261-281
- 中沢道彦 2016「イネ・アワ・キビはいつ来てどのように栽培されたのか」『いま、アッコンが面白い!―タネ・ムシ圧痕が語る先史・古代の農とくらし―』日本学術振興会科学研究費補助金研究成果公開シンポジウム 熊本大学文学部小畑研究室
- (野生種)
-
- タイヌビエ E. oryzicola Vasing. :水田のみに分布
- ヒメタイヌビエ E. crus-galli var. formosensis Ohwi :水田のみに分布
- イヌビエ E. crus-galli var. crus-galli :水田を含む低湿地に分布
- ヒメイヌビエ E. crus-galli var. praticola Ohwi :路傍・畑地を含む乾燥地に分布
- (栽培種)
-
- ヒエ E. utilis
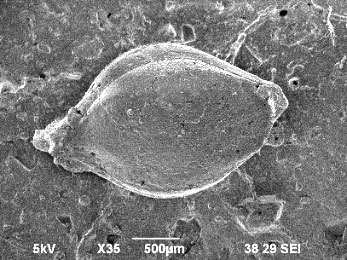 イネ科キビ族ヒエ属に属する草本。穎果はやや菱形の卵形であり、内穎側が平坦で外穎側は逆「く」の字状に大きく膨らむため、横断面形は隅丸の三角形を呈します。最大部は内外穎共に中位にあります。胚部は粒長の約2/3を占めます。反対側の面には尖った臍があります。
イネ科キビ族ヒエ属に属する草本。穎果はやや菱形の卵形であり、内穎側が平坦で外穎側は逆「く」の字状に大きく膨らむため、横断面形は隅丸の三角形を呈します。最大部は内外穎共に中位にあります。胚部は粒長の約2/3を占めます。反対側の面には尖った臍があります。
 ヒエの祖先野生種はイヌビエとされており、北海道・東北地方で縄文時代前期には栽培化されていた可能性が高いです(那須2018)。それらの種子はイヌビエよりも丸みを帯びており、「縄文ヒエ」と名付けられました(吉崎2003)。その後、擦文期になると、イネなどと共に南の地域から東北地方を通り、北海道に別種のヒエが入ってきたと考えられています(小畑2011)。
ヒエの祖先野生種はイヌビエとされており、北海道・東北地方で縄文時代前期には栽培化されていた可能性が高いです(那須2018)。それらの種子はイヌビエよりも丸みを帯びており、「縄文ヒエ」と名付けられました(吉崎2003)。その後、擦文期になると、イネなどと共に南の地域から東北地方を通り、北海道に別種のヒエが入ってきたと考えられています(小畑2011)。

- 参考文献
-
- 小畑弘己 2011「縄文時代の栽培植物とその起源」『東北アジア古民族植物学と縄文農耕』pp.153-179 同成社
- 那須浩郎 2018「縄文時代の植物のドメスティケーション」『第四紀研究』(The Quaternary Research)57(4) pp.109-126
- 吉崎晶一 2003「先史時代の雑穀―ヒエとアズキの考古学―」『雑穀の自然史―その起源と文化を求めて―』pp.52-70 北海道大学図書館刊行会
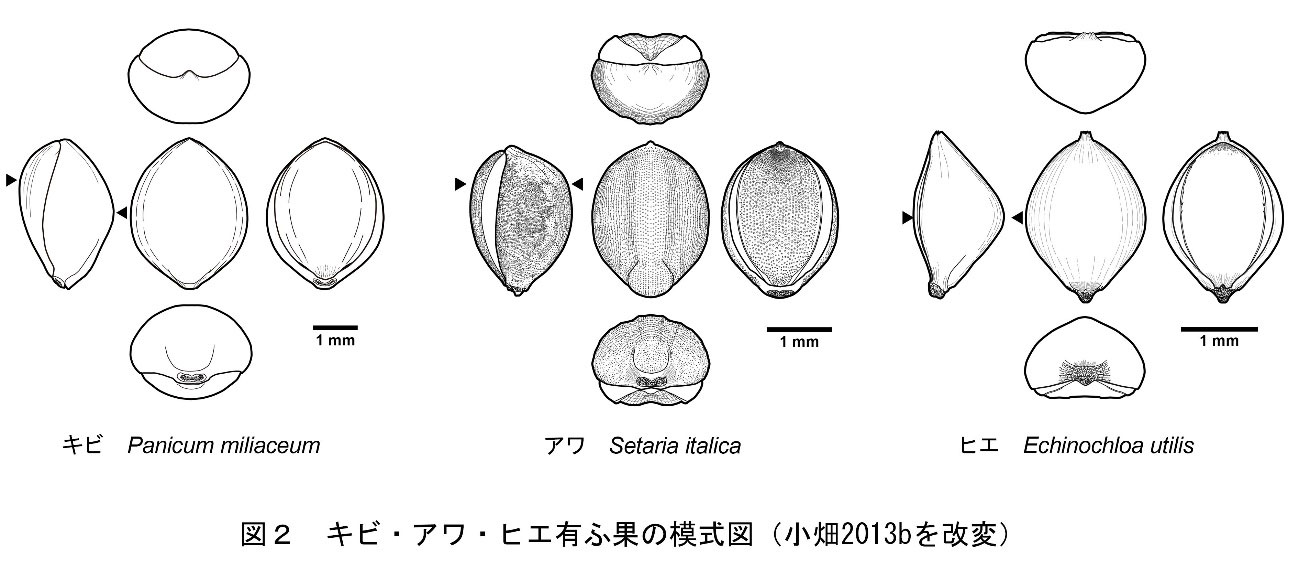
 イネ科キビ亜科キビ族キビ属の一年生草本で、穀物の一種。有粰果は平面観が丸みを帯びた紡錘形を呈し、内外穎の表面は平滑です。外穎の先端は亀の口吻状に突き出します(中山ほか2013)。側面部での最大部は内穎側が上部に、外穎側が中位にあります。内外穎とも膨らんでいるため、横断面形は楕円形を呈しています(小畑・真邉2014)。胚部は粒長の約1/2を占めます。反対側の面にはヘラ形の臍があります(椿坂1993)。
イネ科キビ亜科キビ族キビ属の一年生草本で、穀物の一種。有粰果は平面観が丸みを帯びた紡錘形を呈し、内外穎の表面は平滑です。外穎の先端は亀の口吻状に突き出します(中山ほか2013)。側面部での最大部は内穎側が上部に、外穎側が中位にあります。内外穎とも膨らんでいるため、横断面形は楕円形を呈しています(小畑・真邉2014)。胚部は粒長の約1/2を占めます。反対側の面にはヘラ形の臍があります(椿坂1993)。
 栽培キビの原産地は中央・東アジアの大陸性気候の温帯地域と推定されていますが、原種は不明です。中国では8000年前から栽培されており、主産地の山西省で紀元前6世紀には主作物でした。一方で、日本の古代にはキビの記録が見当たらず、931~936年の和妙類聚抄に「秬黍」とあるのが初出の記録です(小畑2011)。圧痕資料では、島根県三田谷Ⅰ遺跡の約2700年前の例が最古のものです(中沢2016)。
栽培キビの原産地は中央・東アジアの大陸性気候の温帯地域と推定されていますが、原種は不明です。中国では8000年前から栽培されており、主産地の山西省で紀元前6世紀には主作物でした。一方で、日本の古代にはキビの記録が見当たらず、931~936年の和妙類聚抄に「秬黍」とあるのが初出の記録です(小畑2011)。圧痕資料では、島根県三田谷Ⅰ遺跡の約2700年前の例が最古のものです(中沢2016)。

- 参考文献
-
- 小畑弘己 2011「縄文時代の栽培植物とその起源」『東北アジア古民族植物学と縄文農耕』pp.153-179 同成社
- 小畑弘己・真邉 彩 2014「韓国櫛文土器文化の土器圧痕と初期農耕」『国立歴史民俗博物館研究報告』第187集 pp.111-160
- 椿坂恭代 1993「アワ・ヒエ・キビの同定」『先史学と関連科学』吉崎晶一先生還暦記念論集 pp.261-281
- 中沢道彦 2016「イネ・アワ・キビはいつ来てどのように栽培されたのか」『いま、アッコンが面白い!―タネ・ムシ圧痕が語る先史・古代の農とくらし―』日本学術振興会科学研究費補助金研究成果公開シンポジウム 熊本大学文学部小畑研究室
- 中山誠二・庄田慎矢・外山秀一・網倉邦生・兪 炳燮・原田 幹・植月 学 2013「韓国内における雑穀農耕起源の探求」『山梨県立博物館研究紀要』7 pp.1-21
食用マメのうち、遺跡から出土する縄文時代~古墳時代の栽培マメはダイズとアズキに限られ、ササゲは古代(9世紀)以降に、リョクトウは中世(14世紀)に、その他のインゲンやエンドウマメなどは江戸時代~明治時代の初めに日本にもたらされました(小畑2016)。最近、エンドウマメの最古の例(16世紀後半:炭化資料)が山梨県甲府市の武田氏館跡から発見されています(野本ほか2018)。日本最古のマメ科資料の例は、宮崎県王子山遺跡にて検出された約13,000年前の縄文時代草創期のダイズ属圧痕です。縄文時代中期の6~7000年前には中部高地・西関東地方にてダイズ属、ア最近、エンドウマメの最古の例(16世紀後半:炭化資料)が山梨県甲府市の武田氏館跡から発見されています(野本ほか2018)。ズキ亜属が栽培され始めたと考えられています。
- (野生種)
-
- ツルマメ G. m. subsp. soja
- (栽培種)
-
- ダイズ G. m. subsp. max



マメ科マメ亜科ダイズ属の一年生草本。種子は扁平楕円形、もしくは楕円形を呈し、小口側中央に長楕円形の臍を有します。臍の周縁は土手上に盛り上がり、中央部はわずかに凹み、縦溝(Hilar groove)が走ります。臍の下部に種瘤(Lens)はなく、上部には幼根(Radicular lobe)のふくらみが認められます。
また、九州地方を中心に「ワクド石タイプ」という圧痕が検出されています。熊本県ワクド石遺跡で初めて検出され(中沢・丑野2005)、「稲籾」以外の何らかの「植物種子」と考えられていましたが、「ダイズ属のへそ」であることが判明しました(小畑ほか2007)。
- (野生種)
-
- ヤブツルアズキ V. a. var. nipponensis
- (栽培種)
-
- アズキ V. a. var. angularis



マメ科マメ亜科ササゲ属アズキ亜属の一年生草本。種子は両端が平坦な俵形をしており、その側面やや下寄りに方形に近い長楕円形の臍を持ちます。臍の表面はスポンジ状の厚膜が覆っています。臍の下には小さな種瘤が認められます。
- 参考文献
-
- 小畑弘己 2016「ダイズと縄文人」『タネをまく縄文人 最新科学が覆す農耕の起源』pp.8-30 吉川弘文館
- 中沢道彦・丑野毅 2005「レプリカ法による熊本県ワクド石遺跡出土土器の種子状圧痕の観察」『肥後考古』第13号 pp.24-37
- 野本紗英里・佐々木由香・バンダリ スダルシャン 2018「武田氏館跡からみる中世の植物利用」『日本植生史学会第33回大会講演要旨集』,pp. 41-42
 ウルシ科ウルシ属の落葉高木。核は非対称的腎臓形で、長側辺がわずかにくびれます。背腹両面は広線形(偏平)を呈します。ウルシは中国原産の植物とされており、日本列島に野生のウルシは自生していません。漆液を採取し、器に塗って漆器を作るために古くから栽培されています。ウルシの樹液は触るとかぶれるので注意が必要です。
ウルシ科ウルシ属の落葉高木。核は非対称的腎臓形で、長側辺がわずかにくびれます。背腹両面は広線形(偏平)を呈します。ウルシは中国原産の植物とされており、日本列島に野生のウルシは自生していません。漆液を採取し、器に塗って漆器を作るために古くから栽培されています。ウルシの樹液は触るとかぶれるので注意が必要です。
 ウルシの木・果実・花粉や漆器の存在から、縄文時代早期末には日本列島に生育し、前期以降には東日本を中心とした地域の集落周辺に植栽され、漆器製作に用いられたことがわかっています。草創期か早期にウルシと漆液を活用する技術が日本列島の外から伝播したとされています(能城・佐々木2014)。
ウルシの木・果実・花粉や漆器の存在から、縄文時代早期末には日本列島に生育し、前期以降には東日本を中心とした地域の集落周辺に植栽され、漆器製作に用いられたことがわかっています。草創期か早期にウルシと漆液を活用する技術が日本列島の外から伝播したとされています(能城・佐々木2014)。

- 参考文献
-
- 能城修一・佐々木由香 2014「遺跡出土植物遺体からみた縄文時代の森林資源利用」『国立歴史民俗博物館研究報告』第187集 pp.15-48
- シソ Perilla frutescens var. crispa
- エゴマ Perilla frutescens var. frutescens
 シソ科シソ属の一年生草本。果実は広倒卵形で細く低い隆起の大型網目模様が認められ、カモの嘴状の着点部は基部の大部分を占め、中央にわずかな突起があります。
シソ科シソ属の一年生草本。果実は広倒卵形で細く低い隆起の大型網目模様が認められ、カモの嘴状の着点部は基部の大部分を占め、中央にわずかな突起があります。
縄文時代早期から晩期にかけて、九州・北陸・中部・関東地方を中心に分布が認められますが、東北・北海道からは発見されていません。また、時期ごとの出土比率にさほど差はありません。
 シソとエゴマの形態は大変似ており区別がつきにくいですが、シソが比較的小型な果実であるのに対し、エゴマは大型です。これを考慮すると、遺跡出土果実のサイズ比較では、エゴマ型の果実が優位を占めています(中山2018)。休眠性を持つシソは自生的な状態で育成しますが、休眠性を持たないエゴマは人の保護下で安全な時期に播種されなければ、種子が石のように硬くなってしまいます(小畑2011)。つまり、エゴマには人の関与が不可欠であり、縄文時代のエゴマ型果実の存在は、当時の人々によって栽培されていた可能性が高いことを示しています。
シソとエゴマの形態は大変似ており区別がつきにくいですが、シソが比較的小型な果実であるのに対し、エゴマは大型です。これを考慮すると、遺跡出土果実のサイズ比較では、エゴマ型の果実が優位を占めています(中山2018)。休眠性を持つシソは自生的な状態で育成しますが、休眠性を持たないエゴマは人の保護下で安全な時期に播種されなければ、種子が石のように硬くなってしまいます(小畑2011)。つまり、エゴマには人の関与が不可欠であり、縄文時代のエゴマ型果実の存在は、当時の人々によって栽培されていた可能性が高いことを示しています。
- 参考文献
-
- 小畑弘己 2011「オオムギ種子の形態学的観察」『東北アジア古民族植物学と縄文農耕』pp.209-219 同成社
- 中山誠二 2018「栽培植物からみた日本列島の農耕起源―弥生型農耕の系譜―」『境界の考古学』pp.175-184 日本考古学協会2018年度静岡大会実行委員会編
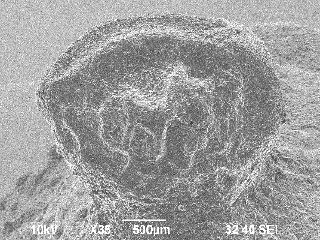 ミカン科サンショウ属の落葉樹。種子の長さは3~4㎜大で、側面観はD字形を呈し、外表面には大きな亀甲状の窪みがあります。広い線形の深い溝状の臍は、腹面の正中線上にあります。
ミカン科サンショウ属の落葉樹。種子の長さは3~4㎜大で、側面観はD字形を呈し、外表面には大きな亀甲状の窪みがあります。広い線形の深い溝状の臍は、腹面の正中線上にあります。
 九州地域の縄文時代を中心に数多く検出されています。現在ではほとんど利用されない植物ですが、カラスザンショウの成分には強い殺虫効果があり、当時の貯蔵食物害虫の防駆虫剤として使用された可能性が指摘されています(真邉・小畑2017)
九州地域の縄文時代を中心に数多く検出されています。現在ではほとんど利用されない植物ですが、カラスザンショウの成分には強い殺虫効果があり、当時の貯蔵食物害虫の防駆虫剤として使用された可能性が指摘されています(真邉・小畑2017)

- 参考文献
-
- 真邉 彩・小畑弘己 2017「産状と成分からみたカラスザンショウ果実の利用法について」『植生史研究』26(1) pp.27-40
 コナラ属( Quercus spp.)、マテバシイ属( Lithocarpus spp.)、シイ属( Castanopsis spp.)のブナ科の果実の総称。一部もしくは全体を殻斗に覆われる堅果。堅果は概ね楕円形であり、先端は尖り花柱となっています。表面の殻は厚く、滑らかです。基部には円い座(へそ)が存在します。
コナラ属( Quercus spp.)、マテバシイ属( Lithocarpus spp.)、シイ属( Castanopsis spp.)のブナ科の果実の総称。一部もしくは全体を殻斗に覆われる堅果。堅果は概ね楕円形であり、先端は尖り花柱となっています。表面の殻は厚く、滑らかです。基部には円い座(へそ)が存在します。
 ドングリ類は縄文時代の全時期を通して様々な遺跡から出土しています。とくに西日本の縄文人はドングリ類を利用することが多く、中でもアクのないイチイガシを盛んに利用していたとされています(佐々木2014)。粒径の大きなドングリの果実は土器の器壁には入りにくいですが、殻の破片は圧痕で多量に検出されています。また、シイ属などの比較的粒径の小さな果実は圧痕で見つかることもあります。
ドングリ類は縄文時代の全時期を通して様々な遺跡から出土しています。とくに西日本の縄文人はドングリ類を利用することが多く、中でもアクのないイチイガシを盛んに利用していたとされています(佐々木2014)。粒径の大きなドングリの果実は土器の器壁には入りにくいですが、殻の破片は圧痕で多量に検出されています。また、シイ属などの比較的粒径の小さな果実は圧痕で見つかることもあります。

- 参考文献
-
- 佐々木由香 2014「縄文人の植物利用―新しい研究法からみえてきたこと」『ここまでわかった!縄文人の植物利用』pp.26-45工藤雄一郎/国立歴史民俗博物館 編
 スイカズラ科(レンプクソウ科)ニワトコ属の落葉低木。核は狭楕円形を呈します。背面は円みがあり、腹面の正中線は鈍稜をなします。側面は半狭楕円形。表面には長軸に直角ないぼ状隆条が並びます。
スイカズラ科(レンプクソウ科)ニワトコ属の落葉低木。核は狭楕円形を呈します。背面は円みがあり、腹面の正中線は鈍稜をなします。側面は半狭楕円形。表面には長軸に直角ないぼ状隆条が並びます。
 ニワトコ属は液果であるため直接食したり、果汁を絞って利用したと推定されています(辻ほか2006)。青森県三内丸山遺跡ではニワトコ属が密集し、他にも様々な液果が伴って出土したことから、醸造の可能性も指摘されています(辻2005)
ニワトコ属は液果であるため直接食したり、果汁を絞って利用したと推定されています(辻ほか2006)。青森県三内丸山遺跡ではニワトコ属が密集し、他にも様々な液果が伴って出土したことから、醸造の可能性も指摘されています(辻2005)
- 参考文献
-
- 辻 圭子・辻誠一郎・南木睦彦 2006「青森県三内丸山遺跡の縄文時代前期から中期の種実遺体群と植物利用」『三内丸山遺跡の生態系史』植生史研究 特別第2号 pp.101-120
- 辻誠一郎 2005「縄文時代における果実酒酒造の可能性」『酒史研究』No.22 pp.21-28
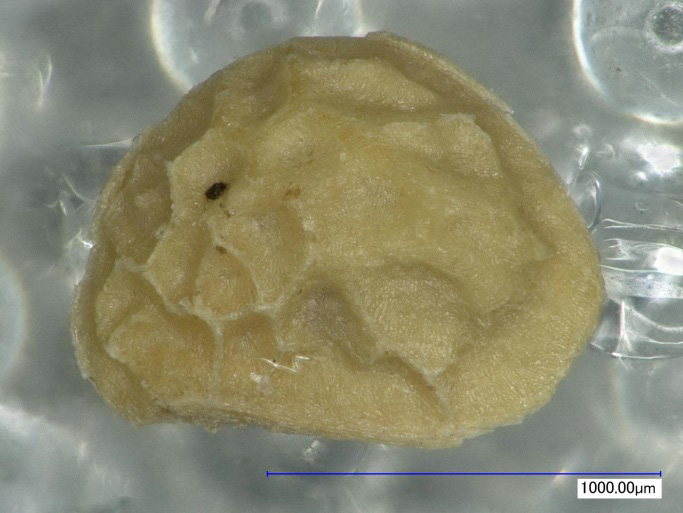
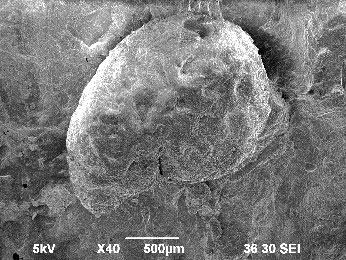 バラ科キイチゴ属の落葉低木。分化が激しく、雑種が非常に多いです。主な種に、クサイチゴ(Rubus hirsutus)、モミジイチゴ(Rubus palmatus var. coptophyllus)、クマイチゴ(Rubus crataegifolius)などがあります。種子は左右非対称の横広卵形を呈し、腹面方向へやや湾曲します。背腹の正中線上に隆条があります。核表面には大型の窪みによる網目模様がみられます。
バラ科キイチゴ属の落葉低木。分化が激しく、雑種が非常に多いです。主な種に、クサイチゴ(Rubus hirsutus)、モミジイチゴ(Rubus palmatus var. coptophyllus)、クマイチゴ(Rubus crataegifolius)などがあります。種子は左右非対称の横広卵形を呈し、腹面方向へやや湾曲します。背腹の正中線上に隆条があります。核表面には大型の窪みによる網目模様がみられます。
 キイチゴ属は日本列島各地でしばしば検出されます。しかし、種子のみでキイチゴ属を区別するのは難しく、種の同定には至っていません。液果であるため、果実を直接食したり、果汁を絞って利用したりした可能性が高いとされています(辻ほか2006)。
キイチゴ属は日本列島各地でしばしば検出されます。しかし、種子のみでキイチゴ属を区別するのは難しく、種の同定には至っていません。液果であるため、果実を直接食したり、果汁を絞って利用したりした可能性が高いとされています(辻ほか2006)。
- 参考文献
-
- 辻 圭子・辻誠一郎・南木睦彦 2006「青森県三内丸山遺跡の縄文時代前期から中期の種実遺体群と植物利用」『三内丸山遺跡の生態系史』植生史研究 特別第2号 pp.101-120
- (ブドウ科ブドウ属)
-
- エビヅル Vitis ficifolia :山野の林縁に生える
- サンカクヅル Vitis fiexuosa:山地に生える
- サンカクヅル Vitis coignetiae:山地に生える
- (ブドウ科ツタ属)
-
- ツタ Parthyenocissus tricuspidata :山野の林縁に生える
- (ブドウ科ノブドウ属)
-
- ノブドウ Ampelopsis glandulosa var. heterophylla :山野の林縁に生える
 果実は有毒。種子は広倒卵形で、基部が短く嘴状に尖っています。側面は狭倒卵形を呈し、背面は円みがあり、倒ヘラ状の窪みが顕著です。
果実は有毒。種子は広倒卵形で、基部が短く嘴状に尖っています。側面は狭倒卵形を呈し、背面は円みがあり、倒ヘラ状の窪みが顕著です。
 ブドウ類は日本列島各地でしばしば検出されていますが、ブドウ類の区別は難しく、現状種の同定には至っていません。液果であるブドウ属は生食したり、果汁を絞って利用したりしたと推定されています(辻ほか2006)。
ブドウ類は日本列島各地でしばしば検出されていますが、ブドウ類の区別は難しく、現状種の同定には至っていません。液果であるブドウ属は生食したり、果汁を絞って利用したりしたと推定されています(辻ほか2006)。

- 参考文献
-
- 辻 圭子・辻誠一郎・南木睦彦 2006「青森県三内丸山遺跡の縄文時代前期から中期の種実遺体群と植物利用」『三内丸山遺跡の生態系史』植生史研究 特別第2号 pp.101-120
- (エゾムギ類)
-
- エゾムギ Elymus sibiricus:寒冷地の山の草原に生える
- ハマムギElymus dahuricus heterophylla :海岸、まれに内陸に生える
- (カモジグサ類)
-
- カモジグサ Elymus tsukushiensis var. transiens:平地の路端、草地に生える
- アオカモジグサ Elymus racemifer :農耕地、市街地の道端に生える
 イネ科エゾムギ属の多年草で、雑草の類。果実は線状長楕円形で、背面は円みがあり、腹面の両側縁は内方にやや曲がり桶状を呈しています。基部の胚部は短い突起状をなします。護衛の先端部は芒(のぎ)へと続きます。エゾムギ・ハマムギ・カモジグサが護衛と内穎の長さが同じなのに対し、アオカモジグサは護衛が内穎よりやや長いです。
イネ科エゾムギ属の多年草で、雑草の類。果実は線状長楕円形で、背面は円みがあり、腹面の両側縁は内方にやや曲がり桶状を呈しています。基部の胚部は短い突起状をなします。護衛の先端部は芒(のぎ)へと続きます。エゾムギ・ハマムギ・カモジグサが護衛と内穎の長さが同じなのに対し、アオカモジグサは護衛が内穎よりやや長いです。
 圧痕でしばしば検出されています。
圧痕でしばしば検出されています。
 ヒユ科イノコヅチ属の多年草。長楕円形の果実は花後閉じた花被片に包まれています。花被には、先が鉤状に曲がった太く長い小胞が二つあり、これで衣服などについて運ばれます。俗にいう「ひっつき虫」です。
ヒユ科イノコヅチ属の多年草。長楕円形の果実は花後閉じた花被片に包まれています。花被には、先が鉤状に曲がった太く長い小胞が二つあり、これで衣服などについて運ばれます。俗にいう「ひっつき虫」です。
 ヌスビトハギ属同様しばしば圧痕でみられ、当時の人々の衣服に張り付いて集落内に持ち込まれたと考えられます。
ヌスビトハギ属同様しばしば圧痕でみられ、当時の人々の衣服に張り付いて集落内に持ち込まれたと考えられます。
 マメ科マメ亜科ヌスビトハギ連ヌスビトハギ亜連ヌスビトハギ属。果実は普通2節あり、各節に1個のD字型の種子が入っています。圧痕では果実が1節になった状態が多いです。果実は半月上で、縁はやや肥厚します。果実の表面にかぎ状の毛が密集した俗にいう「ひっつき虫」の一種です。
マメ科マメ亜科ヌスビトハギ連ヌスビトハギ亜連ヌスビトハギ属。果実は普通2節あり、各節に1個のD字型の種子が入っています。圧痕では果実が1節になった状態が多いです。果実は半月上で、縁はやや肥厚します。果実の表面にかぎ状の毛が密集した俗にいう「ひっつき虫」の一種です。
圧痕ではイノコヅチ同様しばしばみられ、当時の人々の衣服に張り付いて集落内に持ち込まれたと考えられます。
 穎果はやや偏平な狭卵形で、先端が細く尖ります。背面がわずかに曲線をなし、腹面はより湾曲するため、側面観は非対称的な形態です。背面の下部に半円形をした胚があり、腹面の中央には縦溝がみられます。
穎果はやや偏平な狭卵形で、先端が細く尖ります。背面がわずかに曲線をなし、腹面はより湾曲するため、側面観は非対称的な形態です。背面の下部に半円形をした胚があり、腹面の中央には縦溝がみられます。
 ササ属は大群落では数十年に一度しか結実しませんが、小さな集団では毎年結実が確認されており、飢饉や戦時中に食糧として利用されていました。北海道の縄文時代後期から中・近世の遺跡にササ属穎果の炭化種子が出土しており、当時においても結実時には食料資源の一部として利用されていた可能性が指摘されています(山田・椿坂2009)。圧痕では、青森県三内丸山遺跡で縄文時代前期から中期のササ属穎果が検出されています。また、東京都下宅部遺跡では植物珪酸体分析により、縄文時代中期中葉の編組製品にササ属が用いられたことが判明しました(米田・佐々木2014)。
ササ属は大群落では数十年に一度しか結実しませんが、小さな集団では毎年結実が確認されており、飢饉や戦時中に食糧として利用されていました。北海道の縄文時代後期から中・近世の遺跡にササ属穎果の炭化種子が出土しており、当時においても結実時には食料資源の一部として利用されていた可能性が指摘されています(山田・椿坂2009)。圧痕では、青森県三内丸山遺跡で縄文時代前期から中期のササ属穎果が検出されています。また、東京都下宅部遺跡では植物珪酸体分析により、縄文時代中期中葉の編組製品にササ属が用いられたことが判明しました(米田・佐々木2014)。
- 参考文献
-
- 山田悟郎・椿坂恭代 2009「遺跡から出土したササ属種子について」『北海道開拓記念館研究紀要』第37号 pp.13-22
- 米田恭子・佐々木由香 2014「植物珪酸体分析による下宅部遺跡出土網組製品と素材束の素材同定」『国立歴史民俗博物館研究報告』第187集 pp.347-356
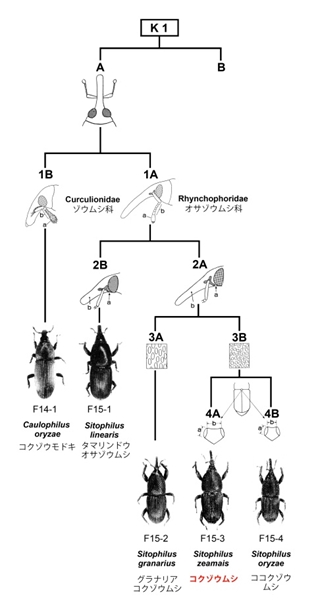
 現在では日本を含む世界共通種のイネやトウモロコシを主に加害する甲虫です。体長は3~4mmであり、長く伸びた口吻が特徴的です。体部全体にほぼ円形の点刻をもち、上翅では複数の点刻列が縦方向の沈線状になります。沈線間の凸部の幅が、点刻による沈線よりも幅が狭いという特徴があります。コクゾウムシとよく似た姉妹種にココクゾウムシShitophilus oryzaeがいます。外面的な特徴はほぼ同じであり、平均体長はコクゾウムシで2.3~3.5mm、ココクゾウムシで2.1~2.9mmと、コクゾウムシの方が若干大きいですが、それぞれ幅があり、重なる部分があります(安富・梅谷1983)。それ以外の決定的な区別点は二箇所あります。コクゾウムシの小楯板の左右両側の隆起部の長さは両者の間隔よりも短いです。ココクゾウの場合は、小楯板の左右両側の隆起部の長さが両者の間隔と同じかより長いです(吉田ほか1989)。また、コクゾウムシとココクゾウムシでは触角の第三節の長さが、コクゾウムシの場合は長さが幅よりも長いが、ココクゾウムシの場合は両者がほぼ等しいという違いがあります(安富・梅谷1983)。
現在では日本を含む世界共通種のイネやトウモロコシを主に加害する甲虫です。体長は3~4mmであり、長く伸びた口吻が特徴的です。体部全体にほぼ円形の点刻をもち、上翅では複数の点刻列が縦方向の沈線状になります。沈線間の凸部の幅が、点刻による沈線よりも幅が狭いという特徴があります。コクゾウムシとよく似た姉妹種にココクゾウムシShitophilus oryzaeがいます。外面的な特徴はほぼ同じであり、平均体長はコクゾウムシで2.3~3.5mm、ココクゾウムシで2.1~2.9mmと、コクゾウムシの方が若干大きいですが、それぞれ幅があり、重なる部分があります(安富・梅谷1983)。それ以外の決定的な区別点は二箇所あります。コクゾウムシの小楯板の左右両側の隆起部の長さは両者の間隔よりも短いです。ココクゾウの場合は、小楯板の左右両側の隆起部の長さが両者の間隔と同じかより長いです(吉田ほか1989)。また、コクゾウムシとココクゾウムシでは触角の第三節の長さが、コクゾウムシの場合は長さが幅よりも長いが、ココクゾウムシの場合は両者がほぼ等しいという違いがあります(安富・梅谷1983)。
ただし、これらの特徴は圧痕レプリカでは観察が困難です。
- 参考文献
-
- 安富和男・梅谷献二1983『原色図鑑 改訂衛生害虫と衣食住の害虫』p.310 全国農村教育協会
- 吉田敏治・渡辺 直・尊田望之 1989『図説貯蔵食品の害虫-実用的識別法から防除法まで-』p.268 全国農村教育協会

 ゴキブリの種類は世界で4000種以上とされていますがその99%が野外に棲息しています。日本で記録されたゴキブリは52種で、その多くが九州以南、特に南西諸島に分布します。圧痕としてゴキブリ成虫本体の発見はありません。
ゴキブリの種類は世界で4000種以上とされていますがその99%が野外に棲息しています。日本で記録されたゴキブリは52種で、その多くが九州以南、特に南西諸島に分布します。圧痕としてゴキブリ成虫本体の発見はありません。
ここでは宮崎県宮崎市本野原遺跡で出土したゴキブリの卵鞘(縄文時代後期前半)について説明します。圧痕は長さ10mm~11mmほどの細長いがまぐち形の形状をもち、縫合線は側辺側で観察できますが、刻みをもつ部分は欠落しています。MNB0488圧痕では縫合線の下部にあたる部分に緩やかな突起が8個並列しています。現生日本の主な屋内種であるヤマトゴキブリ、クロゴキブリ、ワモンゴキブリ、トビイロゴキブリ、チャバネゴキブリと屋外種であるルリゴキブリ、モリチャバネゴキブリ、キョウトゴキブリの卵鞘と比較すると、上記の9種の卵鞘のうち、がまぐち形で縫合線下部に並列する膨らみをもつものは、ヤマトゴキブリ、ワモンゴキブリ、クロゴキブリ、トビイロゴキブリの4種のみです。このうち、全体形が圧痕と類似するのはクロゴキブリとトビイロゴキブリであり、卵の数を比較した場合、MNB0488だと現状では8個しか観察できませんが、復元すると11個になります。圧痕例の推定される卵の数は22個前後であり、12mmに近い卵鞘のサイズの点からもクロゴキブリの卵鞘が圧痕卵鞘にもっとも近いです。よって、本例はクロゴキブリPeriplaneta fuliginosa (Serville)の卵鞘と同定しました。
本野原遺跡から検出されたクロゴキブリの卵鞘圧痕は、クロゴキブリが日本在来種である可能性、もしくは4300年前以前に中国大陸から渡来していた可能性を示すものであり、我が国におけるゴキブリの進化と伝播に関する新たな問題点を提起する発見であるといえます。
- 参考文献
-
- 日本家屋害虫学会 2003『家屋害虫事典』p.468 井上書院
- 小畑弘己 2019『縄文時代の植物利用と家屋害虫-圧痕法のイノベーション』p.258 吉川弘文館
